


ГЕНЕТИЧЕСКИЙ КОД, система "записи" наследств. информации в виде последовательности нуклеотидов в молекулах нуклеиновых к-т. Реализация Г. к. в клетке происходит в два этапа: 1) синтез молекулы матричной, или информационной, РНК (соотв. мРНК, или иРНК) на соответствующем участке ДНК; при этом последовательность нуклеотидов ДНК "переписывается" в нуклеотидную последовательность мРНК (см. Транскрипция); 2)синтез белка, при к-ром последовательность нуклеотидов мРНК переводится в соответствующую последовательность аминокислот (см. Трансляция).
Впервые идея о существовании Г. к. сформулирована А. Дауном и Г. Гамовым в 1952-54, к-рые показали, что последовательность нуклеотидов, однозначно определяющая синтез той или иной аминокислоты, должна содержать не менее трех звеньев. Позднее было доказано, что такая последовательность состоит из трех нуклеотидов, названных кодоном, или триплетом. Т.к. молекулы нуклеиновых к-т, на к-рых происходит синтез мРНК или белка, состоят из остатков только четырех разных нуклеотидов, кодонов, отличающихся между собой, м. б. всего 64.
Все синтезируемые в процессе трансляции белки построены из остатков 20 аминокислот (т. наз. кодируемых). Какой именно кодон ответствен за включение той или иной аминокислоты, можно определить по таблице, в к-рой буквы А, Г, У, Ц обозначают основания, входящие в ну-клеотиды (соотв. аденин, гуанин, урацил, цитозин): в вертикальном ряду слева-в первый нуклеотид кодона, в горизонтальном ряду сверху-во второй, в вертикальном ряду справа-в третий. Трехбуквенные сочетания, напр. фен, сер, лей,-сокращенные назв. аминокислот. Прочерки в таблице означают, что три кодона-УАА, УАГ и УГА в нормальных условиях не кодируют к.-л. аминокислоты. Такие кодоны наз. "бессмысленными", или нонсенс-кодонами. Они являются "сигналами" остановки синтеза полипептидной цепи.
В таблице представлены не все аминокислоты, встречающиеся в белках. В ней нет гидроксипролина и гидроксилизина, содержащихся в коллагене; фосфосерина-компонента всех фосфопротеидов; иодпроизводных тирозина, содержащихся в тиреоглобулине; цистина, к-рый часто встречается в белках, и нек-рых др. аминокислот. Все они-производные др. аминокислот, к-рые кодируются мРНК. Они образуются в результате модификации белков, происходящей после трансляции.
Г.к. специфичен: это означает, что каждый кодон кодирует только одну аминокислоту. Лишь два кодона, кодирующие валин (ГУГ) и метионин (АУГ), способны выполнять дополнит. ф-ции. Если они находятся в начале считываемой области мРНК, к ним присоединяется транспортная РНК (тРНК), несущая формилметионин, к-рый всегда находится в начале строящейся полипептидной цепи, а по завершении синтеза отщепляется целиком или отщепляет формильный остаток, превращаясь в остаток метионина. Т. обр., кодоны ГУГ и АУГ-инициаторы синтеза полипептидной цепи. Если же они не стоят первыми, то не отличаются по ф-циям от др. кодонов.
ТАБЛИЦА ГЕНЕТИЧЕСКОГО КОДА
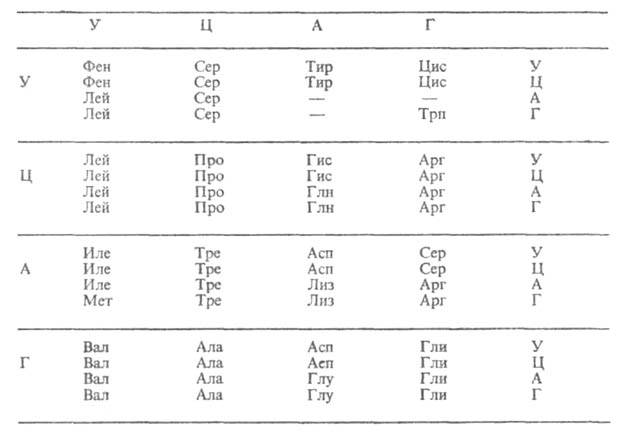
Г.к. называют вырожденным, поскольку 61 кодон кодирует всего 20 аминокислот. Поэтому почти каждой аминокислоте соответствует более чем один кодон. Вырожденность Г.к. неравномерна: для аргинина, серина и лейцина она шестикратна (т.е. для каждой из этих аминокислот имеется по шесть кодонов), тогда как для мн. др. аминокислот (тирозина, гистидина, фенилаланина и др.) лишь двукратна. Две аминокислоты (метионин и триптофан) представлены единств. кодонами. Кодоны-синонимы почти всегда отличаются друг от друга по последнему из трех нуклеотидов, тогда как первые два совпадают. Т. обр., код аминокислоты определяется в осн. первыми двумя "буквами". Вырожденность Г. к. имеет важное значение для повышения устойчивости генетич. информации.
С механизмами трансляции связана еще одна особенность Г.к.: он неперекрывающийся. Кодоны транслируются всегда целиком; для кодирования невозможно использование элементов одного из них в сочетании с элементами соседнего. "Рамкой", ограничивающей транслируемый кодон и перемещающейся скачком сразу на три нуклеотида, служит антикодон тРНК, к-рый представляет собой триплет нуклеотидов, комплементарный одному из кодонов и обусловливающий специфичность к нему. Т. обр., наблюдается линейное соответствие между последовательностью кодирующих триплетов и расположением остатков аминокислот в синтезируемом полипептиде, т.е. код имеет линейный непрерывающийся порядок считывания.
Важнейшее св-во Г. к.-его однонаправленность. Кодоны информативны только в том случае, если они считываются в одном направлении-от первого нуклеотида к последующим.
Г.к. универсален для всех живых существ. Возможны только небольшие видовые изменения, возникшие, вероятно, при эволюции и дифференцировке клеток. Большинство из них связано с вырожденностью кода и проявляется в преимуществ. использовании разных кодонов одной и той же аминокислоты и в различиях в структуре соответствующих тРНК в разных организмах или в разных тканях одного организма.
Представления об общих принципах и осн. св-вах Г. к. были разработаны Ф. Криком в 1957-65. Благодаря работам X. Кораны, X. Маттеи, М. Ниренберга и С. Очоа к 1966 был расшифрован нуклеотидный состав всех кодонов.
Лит.: И час М., Биологический код, пер. с англ., М., 1971. П.
Л. Иванов.