


ЛИПОПОЛИСАХАРИДЫ, смешанные биополимеры, включающие полисахаридную часть и ковалентно связанный с ней липидный остаток.
Л. характерны для микроорганизмов. наиб. подробно изучены Л. грамотрицат. бактерий и, прежде всего, энтеробактерий (содержатся в кишечнике).
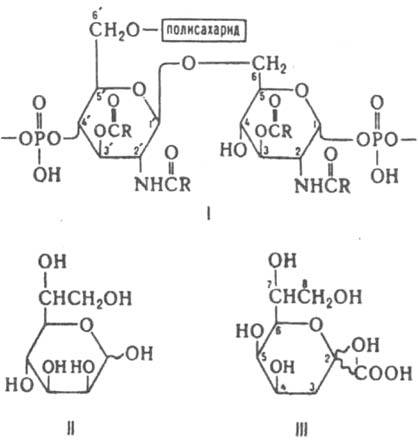
В молекулах Л. гладких штаммов (S-форм) выделяют три различные по структуре участка: липид А, центр. олигосахаридный участок (кор) и О-специфический полисахарид. В шероховатых мутантах (R-формах) полисахаридная цепь отсутствует, а кор м. б. короче, чем в S-формах. Липид А - наиб. консервативная часть Л. и имеет сходную структуру у большинства изученных микроорганизмов. Его основу составляет обычно дисахарид, построенный из двух (b-1 : 6-связанных остатков D-глюкозамина, фосфорилированных в положения 1 и 4' (ф-ла I). Обе аминогруппы и часть гидроксильных групп ацилированы высшими жирными к-тами (до С26), обычно насыщенными и неразветвленными, а также их 3-гидрокси- и 3-ацилоксипроизводными (реже др. производными к-т). Определенный качеств. состав жирных к-т характерен для целых родов и даже семейств бактерий и служит одним из таксономич. критериев. Изменения состава липида А от вида к виду чаще всего связаны с природой и содержанием т. наз. необязательных заместителей (нек-рые моносахариды, остаток 2-аминоэтилфосфорной к-ты), связанных с фосфатными группами. Иногда липидом А м. б. остаток ацилированного моно-, ди- или трисахарида, не содержащий фосфатных групп. Образующие кор остатки моносахаридов группируются в две области: внутреннюю, состоящую из 2-6 остатков высших cахаров (L-глицеро-D-манно-гептозы-II и 3-дезокси-D-манно-октулозоновой к-ты - III), и внешнюю - удаленную от липида А часть молекулы, в к-рую входят остатки гексоз. К остаткам моносахаридов внутр. области присоединяются фосфатные, пирофосфатные и 2-аминоэтилфосфатные группы. Внеш. область кора содержит обычно остатки D-глюкозы, др. сахаров (напр., D-галактозы и D-глюкозамина у энтеробактерий), иногда аминокислот. Кор присоединяется кетозидной связью к одной из неацилированных гидроксильных групп липида А. Эта связь легче гидролизуется к-тами по сравнению с альдозидными связями, находящимися в др. частях молекулы, что позволяет легко разделить углеводный и липидный компоненты Л. хим. путем. С гексозной областью кора связан О-специфический полисахарид. Как правило, он представляет собой регулярный гомо- или гетерополимер, часто разветвленный, построенный из повторяющихся олигосахаридных (от двух до шести остатков моносахаридов) или моносахаридных звеньев. Длина цепи варьирует от одного повторяющегося звена в SR-формах бактерий до 30 и более звеньев в S-формах. Состав полисахаридов чрезвычайно разнообразен. Среди их компонентов насчитываются остатки более 50 разл. моносахаридов (пентоз, гексоз, гексозаминов, дезоксисахаров, уроновых и альдулозоновых к-т, их аминопроизводных, частично метилированных сахаров), а также большое число неуглеводных заместителей (остатков фосфорной к-ты, полиолов, аминов, низших жирных к-т, их гидрокси-, оксо-и аминопроизводных). Структура полисахаридов широко варьирует не только от вида к виду, но и внутри одного вида микроорганизмов. Иногда эти изменения не очень значительны (напр., присоединение к осн. цепи дополнит. остатка моносахарида, О-ацетилирование, замена N-ацильного заместителя на др., изменение конфигурации одного из асимметрич. центров), в др. случаях полностью меняется состав и структура полисахарида. Л. могут быть выделены из клеток экстракцией, напр. р-ром фенола в воде. В водных р-рах молекулы Л. проявляют сильную тенденцию к ассоциации. Образуемые ими агрегаты с мол. массой св. 1 млн. м. б. разрушены путем добавления ПАВ (напр., додецилсульфата Na). Препараты Л. неоднородны, что связано с одновременным присутствием молекул, содержащих и не содержащих полисахаридную цепь, а также в связи с разл. длиной этой цепи. Др. причина неоднородности - присутствие неодинаковых кол-в фосфатных и 2-аминоэтилфосфатных групп в коре, О-ацетильных групп и боковых моносахаридных фрагментов в О-специфическом полисахариде, О-ацильных остатков и необязательных заместителей в липиде А. Л., близкие по строению с Л. грамотрицат. бактерий, продуцируются синезелеными водорослями, в то же время у нек-рых др. микроорганизмов молекулы Л. организованы по-иному. В основе их липидной части находится глицерин или глицерофосфат, к к-рому присоединены высшие жирные к-ты или изопреноидные цепи и полисахаридная часть. У Л. микобактерий (грамположит. палочки, способные к образованию нитчатых форм) липидный остаток отсутствует, а полисахаридная цепь частично ацилирована жирными к-тами (до С8) и янтарной к-той. Биосинтез Л. грамотрицат. бактерий протекает на цитоплазматич. мембране с послед. транспортом макромолекул к месту локализации на внеш. мембране. После образования на начальных стадиях биосинтеза углеводной основы липида А и ацилирования аминогрупп 3-гидроксиалкановыми к-тами происходит перенос остатков 3-дезокси-D-манно-октулозоновой к-ты, а затем негидроксилированных жирных к-т. Далее следует сборка кора путем наращивания цепи на один моносахаридный остаток при участии соответствующих нуклеотидсахаров (в качестве доноров) и специфич. гликозилтрансфераз. Таким же способом на полипренилфосфатном акцепторе строится повторяющееся звено О-специфического полисахарида, к-рое затем подвергается ферментативной полимеризации; образующаяся полимерная цепь при участии фермента транслоказы переносится затем на кор. Альтернативный путь биосинтеза полисахаридной цепи заключается в ее последоват. наращивании на один моносахаридный остаток. Возможны также постполимеризац. модификации полисахарида, такие, как глюкозилирование или О-ацетилирование. Л. играют важную роль в жизнедеятельности бактерий. Они участвуют в транспорте через мембрану разл. соед., являются эндотоксинами, антигенами (т. наз. О-антигенами), а также рецепторами бактериофагов. Липид А, находящийся в тесной ассоциации с протеинами, способствует сохранению целостности и стабильности внеш. мембраны; он же ответствен за токсич. св-ва Л. Полисахаридная цепь ориентирована в сторону окружающей среды, ее тонкая структура определяет узнавание бактерий и специфичность иммунного ответа у высших животных и человека. Лит.: Книрель Ю. А., в кн.: Прогресс химии углеводов, М., 1985, с. 54-76; Wilkinson S. С., в кн.: Surface carbohydrates of the prokaryotic cell, ed. by I. Sutherland, L. - N. Y. - S. F., 1977, p. 97-175; Luderitz O. [a.o.]. "Curr. Top. Membrane Trans.". 1982, v. 17, p. 79-151; Kenne L., Lindberg В., в кн.: The polysaccharides, v. 2. cd. by G.O. Aspinall, N. Y.-L., 1983, p. 287-363: Chemistry of endotoxin, ed. by F.T. Rietschel, Amst., 1984. Ю. А. Книрель.
В молекулах Л. гладких штаммов (S-форм) выделяют три различные по структуре участка: липид А, центр. олигосахаридный участок (кор) и О-специфический полисахарид. В шероховатых мутантах (R-формах) полисахаридная цепь отсутствует, а кор м. б. короче, чем в S-формах. Липид А - наиб. консервативная часть Л. и имеет сходную структуру у большинства изученных микроорганизмов. Его основу составляет обычно дисахарид, построенный из двух (b-1 : 6-связанных остатков D-глюкозамина, фосфорилированных в положения 1 и 4' (ф-ла I). Обе аминогруппы и часть гидроксильных групп ацилированы высшими жирными к-тами (до С26), обычно насыщенными и неразветвленными, а также их 3-гидрокси- и 3-ацилоксипроизводными (реже др. производными к-т). Определенный качеств. состав жирных к-т характерен для целых родов и даже семейств бактерий и служит одним из таксономич. критериев. Изменения состава липида А от вида к виду чаще всего связаны с природой и содержанием т. наз. необязательных заместителей (нек-рые моносахариды, остаток 2-аминоэтилфосфорной к-ты), связанных с фосфатными группами. Иногда липидом А м. б. остаток ацилированного моно-, ди- или трисахарида, не содержащий фосфатных групп. Образующие кор остатки моносахаридов группируются в две области: внутреннюю, состоящую из 2-6 остатков высших cахаров (L-глицеро-D-манно-гептозы-II и 3-дезокси-D-манно-октулозоновой к-ты - III), и внешнюю - удаленную от липида А часть молекулы, в к-рую входят остатки гексоз. К остаткам моносахаридов внутр. области присоединяются фосфатные, пирофосфатные и 2-аминоэтилфосфатные группы. Внеш. область кора содержит обычно остатки D-глюкозы, др. сахаров (напр., D-галактозы и D-глюкозамина у энтеробактерий), иногда аминокислот. Кор присоединяется кетозидной связью к одной из неацилированных гидроксильных групп липида А. Эта связь легче гидролизуется к-тами по сравнению с альдозидными связями, находящимися в др. частях молекулы, что позволяет легко разделить углеводный и липидный компоненты Л. хим. путем. С гексозной областью кора связан О-специфический полисахарид. Как правило, он представляет собой регулярный гомо- или гетерополимер, часто разветвленный, построенный из повторяющихся олигосахаридных (от двух до шести остатков моносахаридов) или моносахаридных звеньев. Длина цепи варьирует от одного повторяющегося звена в SR-формах бактерий до 30 и более звеньев в S-формах. Состав полисахаридов чрезвычайно разнообразен. Среди их компонентов насчитываются остатки более 50 разл. моносахаридов (пентоз, гексоз, гексозаминов, дезоксисахаров, уроновых и альдулозоновых к-т, их аминопроизводных, частично метилированных сахаров), а также большое число неуглеводных заместителей (остатков фосфорной к-ты, полиолов, аминов, низших жирных к-т, их гидрокси-, оксо-и аминопроизводных). Структура полисахаридов широко варьирует не только от вида к виду, но и внутри одного вида микроорганизмов. Иногда эти изменения не очень значительны (напр., присоединение к осн. цепи дополнит. остатка моносахарида, О-ацетилирование, замена N-ацильного заместителя на др., изменение конфигурации одного из асимметрич. центров), в др. случаях полностью меняется состав и структура полисахарида. Л. могут быть выделены из клеток экстракцией, напр. р-ром фенола в воде. В водных р-рах молекулы Л. проявляют сильную тенденцию к ассоциации. Образуемые ими агрегаты с мол. массой св. 1 млн. м. б. разрушены путем добавления ПАВ (напр., додецилсульфата Na). Препараты Л. неоднородны, что связано с одновременным присутствием молекул, содержащих и не содержащих полисахаридную цепь, а также в связи с разл. длиной этой цепи. Др. причина неоднородности - присутствие неодинаковых кол-в фосфатных и 2-аминоэтилфосфатных групп в коре, О-ацетильных групп и боковых моносахаридных фрагментов в О-специфическом полисахариде, О-ацильных остатков и необязательных заместителей в липиде А. Л., близкие по строению с Л. грамотрицат. бактерий, продуцируются синезелеными водорослями, в то же время у нек-рых др. микроорганизмов молекулы Л. организованы по-иному. В основе их липидной части находится глицерин или глицерофосфат, к к-рому присоединены высшие жирные к-ты или изопреноидные цепи и полисахаридная часть. У Л. микобактерий (грамположит. палочки, способные к образованию нитчатых форм) липидный остаток отсутствует, а полисахаридная цепь частично ацилирована жирными к-тами (до С8) и янтарной к-той. Биосинтез Л. грамотрицат. бактерий протекает на цитоплазматич. мембране с послед. транспортом макромолекул к месту локализации на внеш. мембране. После образования на начальных стадиях биосинтеза углеводной основы липида А и ацилирования аминогрупп 3-гидроксиалкановыми к-тами происходит перенос остатков 3-дезокси-D-манно-октулозоновой к-ты, а затем негидроксилированных жирных к-т. Далее следует сборка кора путем наращивания цепи на один моносахаридный остаток при участии соответствующих нуклеотидсахаров (в качестве доноров) и специфич. гликозилтрансфераз. Таким же способом на полипренилфосфатном акцепторе строится повторяющееся звено О-специфического полисахарида, к-рое затем подвергается ферментативной полимеризации; образующаяся полимерная цепь при участии фермента транслоказы переносится затем на кор. Альтернативный путь биосинтеза полисахаридной цепи заключается в ее последоват. наращивании на один моносахаридный остаток. Возможны также постполимеризац. модификации полисахарида, такие, как глюкозилирование или О-ацетилирование. Л. играют важную роль в жизнедеятельности бактерий. Они участвуют в транспорте через мембрану разл. соед., являются эндотоксинами, антигенами (т. наз. О-антигенами), а также рецепторами бактериофагов. Липид А, находящийся в тесной ассоциации с протеинами, способствует сохранению целостности и стабильности внеш. мембраны; он же ответствен за токсич. св-ва Л. Полисахаридная цепь ориентирована в сторону окружающей среды, ее тонкая структура определяет узнавание бактерий и специфичность иммунного ответа у высших животных и человека. Лит.: Книрель Ю. А., в кн.: Прогресс химии углеводов, М., 1985, с. 54-76; Wilkinson S. С., в кн.: Surface carbohydrates of the prokaryotic cell, ed. by I. Sutherland, L. - N. Y. - S. F., 1977, p. 97-175; Luderitz O. [a.o.]. "Curr. Top. Membrane Trans.". 1982, v. 17, p. 79-151; Kenne L., Lindberg В., в кн.: The polysaccharides, v. 2. cd. by G.O. Aspinall, N. Y.-L., 1983, p. 287-363: Chemistry of endotoxin, ed. by F.T. Rietschel, Amst., 1984. Ю. А. Книрель.